Effects of fire on plants and animals: population level
How do populations as a whole fare after fire, or under different fire regimes?
The interaction between life history traits and fire regime can influence the survival of plants and animals at the population scale. Populations refer to a group of individuals within a species that have the potential to interact through space and time.
There is concern about population decline in some mammal species in northern Australia. It is hypothesized that this decline may be due to changed fire regimes. However the evidence is equivocal and there is no obvious explanation as to why certain species populations are in decline. Fire may be part of the story.
Fire type – single or multiple?
There are a variety of different population responses to fires ranging from an increase post-fire, decrease post-fire or no change. An important consideration to make when assessing the effect of fires on biota is that single fire events can have a very different effect to multiple or repeated fires.
The illustration below indicates the difference between a single fire event and multiple fire events, more commonly referred to as a fire regime. You may want to review the discussion of fire regime in the Fire Fundamentals section.
The immediate effects of fire versus longer term effects, need to be identified as populations may behave in different ways in the long and short term. For example, a species may increase in abundance immediately after a single fire, however in the longer term with repeated burning, the species population may decrease.
Furthermore, the effects of a change in regime may not necessarily be clear until some time later. This is illustrated in the following diagram. A new fire regime was implemented at time A, but the effect was not detected until time B.
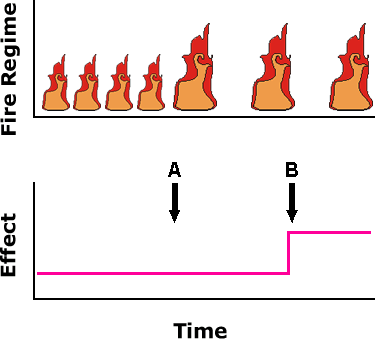
A new fire regime was implemented at time A, but the effect was not detected until time B
Multiple fires
Plants
There are many different hypotheses that have been put forward to explain plant population responses to fire regimes. These include the 'fire interval hypothesis' (constant fire intervals constrain population growth) and ‘event-dependent effects’. It is thought that although fires may be at the same interval different population responses may be produced because intensity (and fire type), season and area burnt can vary independently of frequency (Bond & van Wilgen 1996).
Life-history traits are important because they determine how a population will respond to fire and they can explain what regulates the following processes:
- adult mortality caused by fire
- density of germination post-fire
- establishment of seedlings and subsequent survivorship post-fire
Several authors have attempted to define life-history traits of plants. Noble & Slatyer (1980) developed a ‘vital attribute’ system which can be used to predict population responses to fire. The functional groups are derived based on four processes:
- means of arrival at burnt site
- means of persistence within the site
- pattern of establishment of new species
- three critical time periods (time to reach maturity, time for all plants to die out in a site, & time for all propagules to die out in a site)
Activity: Plant life-history traits & fire responses
Can we classify plant responses to fire?
Gill’s (1981) examination of life-history traits emphasised mode of fire survival (sprouting/non-sprouting) and the type and position of regenerating tissue (seed/bud, above/below ground). Non-sprouters or obligate seeders are woody plants which cannot re-sprout from their trunks. They tend to be killed by fires, and are completely dependent on seed for regeneration.
Note that this classification does not provide information on whether seeding establishment is stimulated by fire.
The plant life history classification is illustrated in Gill (1981).
Reading:
Gill, A.M. (1981) Adaptive responses of Australian vascular plant species to fires. In: Fire and the Australian biota (Gill, A.M., Groves, R.H. & Noble, I.R., eds.), p.582. Australian Academy of Science, Canberra.
or
Bond, W.J. & van Wilgen, B.W. (1996) Fire and Plants, p.47. Chapman & Hall, London.
Further reading
Another classification of fire life-histories is by Bond & van Wilgen (1996), and is based on seedling recruitment (stimulation by fire, or none) and vegetation survival (sprouters/non-sprouters).
Reading:
Bond, W.J. & van Wilgen, B.W. (1996) Fire and Plants, p.50. Chapman & Hall, London.
Activity: Plant life-history & survival
Why does life-history matter?
Life-history strategy can play an important role in influencing the survival and persistence of species. Williams et al. (2002) illustrate how survival in a given environment can depend on the life-history strategy of the species concerned.
Reading:
Williams, R.J., Griffiths, A.D. & Allan, G.E. (2002) Fire regimes and biodiversity in the savannas of northern Australia. In: Flammable Australia (Bradstock, R.A., Williams, J.E. & Gill, A.M., eds.), pp. 289-293. Cambridge University Press, Cambridge.
Activity: Obligate seeders & fire regime
Will obligate seeders survive changing fire regimes?
Obligate seeders are plants that can only regenerate after fire from seed. If fires are too frequent plants cannot reach maturity to set seed and species persistence may be threatened. Russell-Smith et al. (2002) look at obligate seeders and the potential risk associated with this strategy given changing fire regimes across much of Arnhemland in northern Australia.
Russell-Smith, J., Ryan, P.G. & Cheal, D. (2002) Fire regimes and the conservation of sandstone heath in monsoonal northern Australia: frequency, interval, patchiness. Biological Conservation 104: 91-106.
Further reading
Zammit & Westoby (1988) report on the link between complete serotiny and life-history trait (re-sprouter or obligate seeder).
Reading:
Zammit, C. & Westoby, M. (1988) Pre-dispersal seed losses and the survival of seeds and seedlings of two serotinous Banksia shrubs in burnt and unburnt heath. Journal of Ecology 76: 200-214.
Activity: Fire regime & tree survival
Which trees will survive?
The effect of fire intensity and season of burn on the survival of biota depends on their life-history.
This paper considers how the prevailing fire regime influences fire intensities which in turn affects tree survival.
How do you think sprouters and non-sprouters might be affected by intensity and season of burn, and what influence would plant size have? Williams et al. (1998) discuss this.
Reading:
Williams, R.J., Cook, G.D., Gill, A.M. & Moore, P.H.R. (1998) Fire regime, intensity and tree survival in a tropical savanna in northern Australia. Australian Journal of Ecology 24: 50-59.
Activity: Eucalyptus seedling survival & fire
How do seedlings establish in relation to fire?
Fire frequency is also important because it can affect plant and animal survival.
This paper focuses on Eucalyptus seedling establishment in relation to fires at the Kapalga fire experiment in Kakadu National Park.
How does fire frequency affect seedling survival and what does the author suggest this might mean for fire management?
Reading:
Setterfield, S.A. (2002) Seedling establishment in an Australian tropical savanna: effects of seed supply, soil disturbance and fire. Journal of Applied Ecology 39: 949-959.
Animals
Life-history traits in animals, in the context of fire responses, have received far less attention. Traits that have been suggested include the ability to find refugia (local mobility in face of fire), long-distance dispersal ability, breadth of diet (or ability to switch diet after fire), susceptibility to predation and competition, habitat specificity, and intrinsic rate of population increase after fire.
Further reading
Many faunal life-history traits and responses to fire will be the same across the country, from northern Australia to the mallee and heath vegetation of the south.
This excellent article clearly demonstrates the ways in which life-history traits vary at different intervals following a fire. Look for similarities and differences for reptiles and small mammals.
However, bear in mind that fires in the mallee system are much less frequent and the consequences of fire can be quite different than in northern savannas.
Reading:
Friend, G. R. (1993) Impact of fire on small vertebrates in mallee woodlands and heathlands of temperate Australia: a review. Biological Conservation 65: 99-114.
Activity: Bandicoot survival & fire frequency
What determines bandicoot survival?
In terms of animals, the following paper reports on a study that found that fire frequency was the most important determinant of survival for the northern brown bandicoot.
How important are fire frequency and season for the survival and persistence of this species? What might this mean for conservation recommendations?
Reading:
Pardon, G. L., Brook, B.W., Griffiths, A.D. & Braithwaite, R.W. (2003) Determinants of survival for the northern brown bandicoot under a landscape-scale fire experiment. Journal of Animal Ecology 72: 106-115.
Declining species in the Top End – is it fire related?
There is mounting concern over the amount of burning in northern Australia. Each year more than 50% of the Top End burns. Burning is thought to be too frequent with many areas of the Top End being burnt annually. It is thought this fire pattern may be having a detrimental effect on biodiversity.
While fire is frequently cited as the major cause for these declines, there may be many other factors contributing to these declines. These include feral animals, weeds, land clearing and land degradation. It is likely that some of these will act in combination to affect biodiversity.
Activity: Investigating declining species
Who is in decline and why?
Look for papers to find out what taxa or species are thought to be in decline. Carefully consider the discussion and conclusions drawn: what evidence is there for fire as the principle cause?
How much of the evidence is speculative (i.e. not based on firm evidence)? Can you think of how a study might be designed to address some of these issues?
Use the reference to list to guide your reading (pdf document)
Further reading
A fire sensitive species: the cypress pine, Callitris intratropica
The native cypress-pine is thought to be in decline across the Arnhemland Plateau in the Top End because of extensive, late-season fires. Fires in October/November are particularly intense, and when they result in 100% foliage burn can kill cypress pines. The trees are slow-growing (minimum of 10 years to mature and produce seeds) which means they are threatened by frequent fires.
Cypress Pine, Callitris intratropica, at the Territory Wildlife Park
For further information on the cypress-pine read Bowman & Panton (1993).
Reading:
Bowman, D.M.J.S. & Panton, W.J. (1993) Decline of Callitris intratropica R.T. Baker & H.G. Smith in the Northern Territory: implications for pre- and post-European colonisation fire regimes. Journal of Biogeography 20: 373-381.

![]()